
Il rapporto Redfield: gli studi accademici

Qua di seguito sono esposti, senza alcuna interpretazione, stralci degli studi effettuati da Bongakov e Levich presso il Dipartimento di Biologia dell’Università Statale di Mosca.
La natura del riarrangiamento della comunità del fitoplancton causato da una variazione del rapporto N/P è mostrata sia per i microcosmi artificiali di laboratorio che per le algocenenosi naturali in vitro e in situ.
Gli esperimenti hanno dimostrato che alti rapporti ponderali N/P (da 20 a 50) favoriscono lo sviluppo di Chlorococcales, mentre un abbassamento del rapporto N/P a valori di 5÷10 porta a una comunità dominata da Cyanophyta. Come dimostrato dalla modellizzazione delle variazioni, l’abbondanza relativa delle specie di fitoplancton dovrebbe dipendere solo dalle quantità relative di N e P nell’ambiente, in modo che il rapporto ottimale N/P per una data specie sia uguale al rapporto tra i requisiti minimi di cellule per questi elementi.
Una prova empirica di questa legge risiede nel fatto che per alcune specie di Clorococcales e Cyanophyta i rapporti dei requisiti per N e P calcolati nell’esperimento erano vicini ai loro rapporti di concentrazione ambientale ottimali. Ad esempio, un aumento di N/P dal valore di 4/1 a 25÷50/1 nell’acqua degli stagni di allevamento del pesce ha portato ad un aumento dell’abbondanza di Chlorococcales, dovuto principalmente allo Scenedesmus quadricauda , che ha all’incirca lo stesso rapporto tra i requisiti. Il rapporto N/P può essere riconosciuto come un fattore ecologico indipendente, distinto dalle concentrazioni di azoto e fosforo prese separatamente.
Introduzione
I macronutrienti primari che limitano lo sviluppo della biomassa di fitoplancton in natura sono il fosforo, l’azoto ed il silicio, quest’ultimo contenuto nella composizione delle alghe diatomee perché è un componente indispensabile per lo sviluppo del frustolo siliceo. Ciascuno di questi elementi biogenici, presi separatamente, possono essere un fattore limitante, come è stato ripetutamente verificato in letteratura. Si è prestata molta meno attenzione alle interazioni tra N, P e Si negli ecosistemi e l’influenza di queste interazioni sulla struttura dell’algocenosi. Tuttavia c’è un gran numero di opere che dimostrano un ruolo significativo di regolazione dei rapporti di concentrazione di N, P e Si in ambiente acquatico (Pearsall, 1930, 1932, Rhee, 1978; Smith, 1983, 1986; Pick and Lean, 1987; Stockner and Shortreed, 1988; Suttle et al., 1991; ecc.). Questi lavori hanno rivelato la natura della ristrutturazione dell’algocenosi derivante dai cambiamenti in questi rapporti sia per esperimenti naturali che di laboratorio con esperimenti su comunità microalgali artificiali.
Sulla base dei loro studi sul problema, gli autori discutono la questione di quale dei fattori determina la composizione di un’algocenosi in misura maggiore, se le concentrazioni degli elementi assoluti o le loro quantità relative.
Microcosmi di laboratorio – dati da studi precedenti
Una quantità significativa di documenti presenta i risultati degli esperimenti su colture sperimentali in laboratorio. Questi esperimenti avevano il seguente schema comune, è stata creata una sequenza variata di rapporti di elementi biogenici aggiungendo al mezzo sostanze diverse, sono stati così trovati i rapporti ottimali.
Tilman (1977) ha studiato una coltura mista semi continue di diatomee A. formosa e Cyclotella meneghiniana Kutz sotto una vasta gamma di rapporti Si/P, quando il rapporto molare di questi elementi biogenici era inferiore a 6/1, A. formosa dominava, mentre quando il rapporto superava 90, dominava C. meneghiniana.
Rhee (1978) ha concluso sulla base dei suoi esperimenti con monoculture di Scenedesmus sp. che per questa alga il rapporto N/P uguale a 30/1 è quello che stimola la crescita, a rapportI più alti e più bassi la crescita della coltura è limitata rispettivamente dall’azoto e dal fosforo. Il rapporto ottimale N/P trovato per il cianobatterio Anacystis nidulans P. Richt. è anche 30/1 (Sirenko, 1972).
Holm e Armstrong (1981) hanno sviluppato una policoltura di due alghe, la diatomea Asterionella formosa Hass. ed il cianobatterio Microcystis aeruginosa Kutz nel regime di coltivazione continua. Nel corso dell’esperimento il rapporto atomico Si/P variava da 2 a 200. All’aumentare del rapporto Si/P , il rapporto di biomassa delle due specie è passato da 1/99 a 96/4. Si è scoperto che A. formosa è una specie con una maggiore domanda di silicio ma ottiene un vantaggio rispetto al rivale nella condizione di carenza di fosforo. È interessante notare che simili rapporti di biomassa si sono formati a diverse concentrazioni assolute di silicio e fosforo ma quando i loro rapporti nel mezzo erano simili. Ad esempio, l’alga M. aeruginosa potrebbe prendere il sopravvento con un rapporto di 25/1 (98% della biomassa totale) e con un rapporto di 100/10 (97%), mentre A. formosa prende il sopravvento a 300/2 ( 93%) e 100/0.5 (96%).
A livello di monocoltura, è stata dimostrata anche una varietà di responsabilità delle specie sotto limitanti variazioni di sostanze. Quando furono confrontati la produttività di due specie di alghe verdi, Scenedesmus quadricauda e Stigeoclonium tenue (Ag.) Kutz, I valori critici del rapporto N/P risultarono essere pari a 22/1 per il primo e 17/1 per il secondo ( Vries e Klapwijk, 1987). Il valore critico è il valore del rapporto più basso che, essendo superato, porta alla limitazione della crescita del fosforo.
La crescita del cianobatterio Synechococcus Nag. in una policoltura con il verde Scenedesmus quadricauda (Turp.) Breb è stato studiato in condizioni di carenza di fosforo, cioè a rapporti N/P elevati (Suttle e Harrison, 1988), e Synechococcus soppresse l’alga verde in competizione.
In esperimenti con una coltura di due alghe marine Skeletonema costatum (Grev.) Cl. e Phaeodactylum tricornutum Bohlin, è stato studiato l’impatto di temperatura, luce e condizioni biogeniche, S. costatum soppiantò Ph. tricornutum a bassi rapporti N/Si e alti rapporti N/P nel mezzo (de Pauw e Naessens-Foucquaert, 1991). Un rapporto ottimale per S. costatum si è rivelato essere N/Si/P = 25/25/1.
Microcosmi di laboratorio – dati da nuovi esperimenti
Nei nostri esperimenti (Levich e Bulgakov, 1993) abbiamo studiato i rapporti ottimali tra azoto e fosforo negli ambienti nutritivi coltivando alcune specie di microalghe in policultura di laboratorio. Abbiamo cercato di controllare la composizione delle specie di una algocenosi artificiale più semplice variando il rapporto iniziale di concentrazione tra azoto e fosforo.
Cloni di laboratorio di microalghe sono stati ottenuti dalla collezione dell’Istituto di Fisiologia Vegetale, dell’Accademia Russa delle Scienze e dalla collezione del Dipartimento di Fisiologia Vegetale dell’Università Statale di Mosca.
Le comunità di alghe di laboratorio utilizzate in questo esperimento sono state coltivate in terreno contenente diverse combinazioni di concentrazioni di N (KNO3) e P (K2HPO4).
Cloni di raccolta puri di alghe sono stati inoculati in fiasche con terreno liquido sterile in condizioni sterili, le policolture ottenute sono state coltivate in un luminostato a illuminazione costante di 7,3 Wt/m2.
Gli esperimenti furono continuati fino a quando tutti i cloni raggiunsero la fase stazionaria di crescita. Il numero di cellule e il peso individuale delle cellule, per mezzo del quale il numero è stato convertito in biomassa, sono stati contati periodicamente al microscopio, contemporaneamente sono state determinate le concentrazioni di elementi nutritivi nel mezzo. La concentrazione di N è stata stimata utilizzando la misurazione degli elettrodi ionoselettivi adattata alle sospensioni microalgali (Bulgakov et al., 1985), la concentrazione di P è stata misurata con un metodo colorimetrico modificato (Rinkis e Nollendorf 1982). Le quote iniziali e finali di cellule N e P sono state stimate per tutte le specie dal metodo di stima del fabbisogno di fitoplancton in nutrienti biogenici in diverse fasi della loro crescita (Levich 1989). Per stimare la quota di biomassa finale di ogni specie cresciuta con le risorse minerali del terreno, l’incremento della biomassa di N e P situato nelle cellule all’inizio dell’esperimento è stato stimato calcolando le quote di cellule iniziali per tutte le specie. La competizione è stata stimata dalla biomassa di ciascuna specie nella fase stazionaria della sua crescita.
Gli esperimenti sono stati condotti con colture microalgali discontinue. I risultati della crescita delle specie in un’algocenosi artificiale sono stati stimati dai loro contributi parziali all’abbondanza totale o alla biomassa della comunità quando la fase stazionaria della crescita era stata raggiunta da tutte le specie della policoltura.
Quattro specie di Clorococcales hanno partecipato alla prima serie di esperimenti: Scenedesmus quadricauda , Chlorella vulgaris Beyer, Ankistrodesmus falcatus (Corda) Ralfs e Ankistrodesmus sp.
Le policolture di queste quattro specie sono cresciute in due terreni di coltura con le seguenti concentrazioni iniziali di N/P: media 0.1÷1.1 mg/l di azoto e 0.3 mg/l di fosforo (N/P di massa = 3,5); mezzo 0.2÷50 mg/l di azoto e 0.25 mg/l di fosforo (N/P = 20).
I 37 giorni dell’esperimento si sono rivelati insufficienti per tutte le specie per raggiungere le loro fasi stazionarie, tuttavia, questa circostanza non nascondeva gli effetti rivelati che sarebbero diventati ancora più chiari se la fase stazionaria fosse stata raggiunta. In entrambi i media, mentre la crescita si verificava sia con le risorse ambientali che con lo stoccaggio delle celle, la comunità era dominata da S. quadricauda. Tuttavia, mentre a N/P = 3,5 questa dominazione non era così schiacciante (44% dell’abbondanza totale), quando il rapporto N/P aumentava fino a 20, la cultura di S. quadricauda praticamente soppiantò le altre tre specie dalla comunità (Figura 1A), la loro abbondanza relativa sommata era pari al 17%. Nel mezzo con N/P = 3.5 solo S. quadricauda e A. il falcatus raggiunse la sua fase stazionaria, mentre a N/P = 20, al contrario, S. quadricauda fu l’unica specie che continuò a crescere alla fine dell’esperimento.
Quindi, portare la crescita di una policoltura fino allo stadio stazionario aumenterebbe molto probabilmente la tendenza osservata. Un confronto tra l’abbondanza relativa di specie coltivate esclusivamente con l’azoto ambientale e il fosforo ha mostrato che una transizione dal rapporto N/P basso a quello alto determina un cambiamento nelle specie dominanti della comunità.
Mentre nella prima serie, in N/P = 3.5, Ch. vulgaris è assolutamente dominante (100%), in N/P = 20, a differenza di quello, S. quadricauda ammonta al 93% dell’intera biomassa. (Figura 1B)

Nella seconda serie di esperimenti la comunità artificiale consisteva di sole due specie di Clorococcales: Scenedesmus quadricauda e Ankistrodesmus falcatus , mentre il rapporto N/P era variato maggiormente: medio 0.1÷0.4 mg/l di azoto e 0.31 mg/l di fosforo (N/P = 1,3); media 0.2÷1.4 mg/l di azoto e 0.31 mg/l di fosforo (N/P = 4,5); medio 0.3÷3.4 mg/l di azoto e 0.06 mg/l di fosforo (N/P = 57).
La crescita della coltura indotta da stoccaggio era modesta rispetto alla crescita della biomassa, quindi le correzioni corrispondenti non erano in grado di alterare sostanzialmente la distribuzione della biomassa finale delle specie in tutti e tre i campioni.
Quando le colture sono state seminate in tutti i campioni, la biomassa iniziale di policoltura consisteva nel 53% di S. quadricauda e del 47% di A. falcatus. All’aumentare del rapporto N/P, S. quadricauda aumentò la sua biomassa relativa dal 78% con N/P = 1,3, al 87% con N/P = 3,7; e al 90,5% con N/P = 57 in massa. La crescita delle scorte cellulari è stata modesta rispetto alla crescita totale della biomassa, pertanto gli emendamenti corrispondenti non hanno comportato modifiche significative nella distribuzione della biomassa finale in tutti e tre i campioni.
Fitoplancton naturale in vitro – dati da studi precedenti
Esperimenti di laboratorio sono stati condotti anche su consorzi di fitoplancton naturale, piuttosto che su miscele artificiali di cloni di laboratorio di alghe. Diventa possibile analizzare l’influenza dei rapporti di elementi biogenici non solo su specie separate di fitoplancton, ma anche su interi gruppi di specie, unificati sia da un marcatore sistematico, sia dalla dimensione. In questo modo le condizioni stabili in laboratorio consentono un controllo severo dei fattori ambientali assegnati.
Sommer (1983), variando il rapporto molare ambientale Si/P da 4 a 30, su popolazioni multi specie coltivate naturali (oltre 30 specie) di fitoplancton lacustre. In condizioni carenti di silicio Si/P=4/1 l’alga Mougeotia thylespora ha dominato. A rapporti approssimativamente equilibrati 10÷40/1 i più numerosi erano Koliella spiculiformis Hind , Synedra acus Kutz. e Asterionella formosa. A carenza di fosforo Si/P=80/1 solo S. l’acus era dominante, l’autore ha osservato questi risultati indipendentemente dalla composizione naturale dell’inoculo. Un confronto con l’abbondanza di specie di fitoplancton ha mostrato che quando i valori dei rapporti degli elementi biogenici erano vicini a quelli sperimentali, i risultati della competizione nel bacino naturale erano simili.
Suttle e Harrison (1988) hanno condotto una serie di esperimenti con fitoplancton naturale d’acqua dolce in condizioni di laboratorio a rapporti N/P=5, 15 e 45.
Con N/P=45 è stata osservata una dominazione assoluta del ciano batterio Sinechococco, con rapporti più bassi i più numerosi erano due diatomee, Nitzschia holsatica Hust. e Synedra radians (Kutz.) Hust., insieme all’alga verde Scenedesmus sp.
Kilham (1986), analizzando la crescita del fitoplancton dal lago Michigan in laboratorio a diversi rapporti ambientali Si/P, ha concluso che è il rapporto di fornitura delle risorse piuttosto che la quantità assoluta che ha determinato l’esito della competizione per le specie. Mostrò che sotto la limitazione del fosforo le diatomee soppiantano i rappresentanti di tutte gli altri gruppi tassonomici di una comunità; tuttavia, con concentrazione di silicio ridotte la dominazione si è spostata sulle alghe verdi. In media con un elevato rapporto Si/P (313 e 74) le diatomee erano dominanti e quelle verdi lo erano a bassi rapporti Si/P (4,6 e 0,9).
I cianobatteri sono stati eliminati in maniera competitiva dall’esperimento poiché erano stati utilizzati rapporti di alimentazione N/P molto alti (100).
Quando il mezzo veniva fornito con Si e P in un rapporto di 20: 1 in moli, le alghe verdi dominavano, mentre le diatomee dominavano a Si: P = 80: 1 (Grover 1989).
Grover (1989), che coltivava una comunità di alghe lacustri usando una coltura semicontinua, arrivò a conclusioni simili. Quando il mezzo era alimentato con silicio e fosforo in un rapporto era di 20, le alghe verdi dominavano, mentre a Si/P=80 predominavano le diatomee.
Quando in una comunità algale di un lago acido è stato inoculato il cianobatterio Anabaena variabilis Kutz. in sacchi di polietilene, un aumento del pH e una diminuzione del rapporto N/P nell’ambiente hanno provocato una crescita della biomassa di fitoplancton senza cambiamento di dominazione ( Wilcox e De Costa, 1990), è stato dimostrato che i valori del rapporto N/P>25 sono alla crescita dei cianobatteri.
Un lago oligotrofico è stato alimentato da fertilizzanti minerali con il rapporto atomico N/P=50, successivamente sono stati prelevati campioni e sono state studiate in laboratorio le cinetiche di consumo N e P di frazioni di due dimensioni (più piccole e più grandi di 3 micrometri), il rapporto N/P consumato è stato calcolato come il rapporto tra i tassi massimi di consumo di questi elementi. Si è scoperto che la frazione di piccole dimensioni (principalmente Synechococcus spp.) è limitata dall’azoto, mentre quella grande (principalmente Rhizosolenia spp. e Cyclotella spp.) dal fosforo, quindi per specie di taglia piccola un rapporto N/P ottimale è superiore a 50, mentre per quelli di taglia grande è inferiore a 50 (Suttle et al., 1991).
Fitoplancton naturale in vitro – dati da nuovi esperimenti
Abbiamo condotto esperimenti di coltura discontinua volti a studiare l’impatto di diversi rapporti di N e P sulla specie e la struttura dimensionale di un’algocenosi prelevata da un laghetto di allevamento ittico (Levich et al., 1997) nella regione di Astrakhan, l’acqua è stata collocata in fiasche da 2 litri e sono state fornite NH4 – NO3 e Ca(H2PO4)2 in diverse combinazioni quantitative, sono stati ottenuti complessivamente cinque valori del rapporto N/P, con la base naturale presa in considerazione: 2/1, 5/1, 20/1, 50/1, 100/1, la biomassa di fitoplancton originario dello stagno era la stessa in tutte le fiasche.
Per eliminare l’effetto del pascolo dello zooplancton, l’acqua da studiare prima dell’esperimento era stata fatta passare attraverso una rete a maglie con dimensioni corrispondenti alla dimensione minima dello zooplancton e quindi collocato nell’oscurità per due giorni, una delle analisi era quella di controllo, cioè la concentrazione di elementi biogenici originali era uguale a quella di fondo. Nel corso dell’esperimento tutte le prove sono state tenute all’aperto a 50 metri dallo stagno utilizzato per il campionamento dell’acqua con condizioni di illuminazione e temperatura corrispondenti a quelle naturali, le osservazioni delle dinamiche di crescita delle alghe sono state condotte entro 8 giorni, la verifica della concentrazione di fitoplancton e dei componenti biogenici della nutrizione minerale venivano effettuate al microscopio. L’analisi della struttura delle dimensioni includeva un confronto delle frazioni di alcune classi dimensionali di alghe nella biomassa.
A tale scopo tutte le specie trovate sono state divise in sei gruppi di dimensioni in base al loro volume: (1) meno di 0,1 ng; (2) da 0,1 a 0,3 (3) da 0,3 a 1 ng; (4) da 1 a 3,2 ng; (5) da 3,2 a 10 ng; (6) più di 10 ng. Le dimensioni effettive delle celle corrispondenti ai loro volumi sono le seguenti: (1) < 5,8 mm, (2) 5,8÷8,3 mm, (3) 8,3÷12,4 mm, (4) 12,4÷18,3 mm, (5) 18,3÷26,7 mm, (6)> 26,7 mm. Con una dimensione efficace si intende il diametro di una cella sferoidale avente una massa identica.
La biomassa finale è stata analizzata sia per i taxon di fitoplancton (Chlorococcales, Chlorophyta, Bacillariophyta e Cyanophyta) sia a livello di generi e specie. I taxon a livello di generi e specie erano divisi in dominanti, la cui biomassa non era inferiore al 20% dell’intera biomassa al sesto giorno dell’esperimento almeno in una singola fiasca, non rappresentativi con una biomassa inferiore a 1% e sottodominanti tutto il resto. Sono state anche confrontate le biomasse delle classi di dimensioni delle alghe.
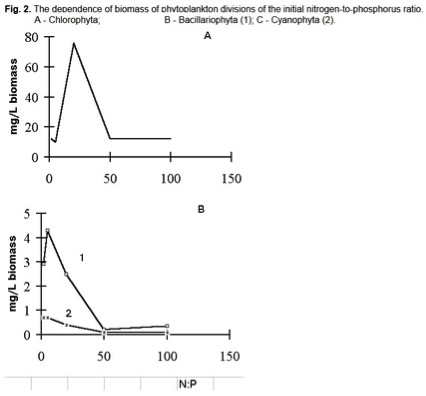
Rapporti di N/P>5 hanno notevolmente trasformato la struttura di algocenosi in direzione di una dominazione assoluta delle verdi Chlorophyta (Figura 2), la loro biomassa aveva un picco unico corrispondente alla crescita più intensa (Figura 2), questo picco è osservato a N/P=20.
Tuttavia, la biomassa delle sue specie predominate Scenedesmus quadricauda presentava un picco a N/P=20 e diminuiva a N/P=50÷100.
La biomassa massima di diatomee e cianobatteri è stata rilevata a bassi rapporti N/P=2÷5 (Figura 2).
I rapporti N/P=5÷20 erano ottimali per la crescita delle diatomee di Stephanodiscus e Nitzschia . Il cianobatterio Microcystis si è sviluppato meglio a rapporti tra 2 e 5. Rapporti N/P più alti hanno dato una minore biomassa tra la Cyanophyta. Al contrario, la biomassa di Euglenophyta, dopo la diminuzione al rapporto N/P=20, è aumentata di nuovo a rapporti N/P=50÷100.
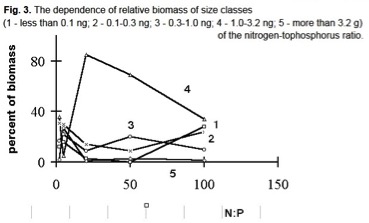
L’abbondanza delle classi di dimensione delle alghe in% della biomassa totale è mostrata in Figura 3, le cellule >10 ng erano estremamente rare nella biomassa e sono escluse. Le cellule dell’intervallo 1÷3,2 ng occupavano una posizione predominante nella comunità con rapporti N/P=20÷50, mentre la loro frazione di biomassa diminuiva a rapporti N/P più alti e più bassi.
Le cellule con massa compresa tra 0,3÷1 ng erano più abbondanti con un rapporto di N/P=5.
La rappresentazione delle due classi più piccole era molto bassa con N/P=2÷50, tuttavia, con un rapporto di 100 ripristinano la loro posizione predominante (Figura 3).
Fitoplancton naturale in situ – dati da studi precedenti
Già negli anni Trenta del secolo attuale Pearsall (1930, 1932) fu uno dei primi ad osservare che i diversi rapporti di abbondanza di elementi chimici, tra cui N/P e Ca+Mg/Na+K, promuovono la formazione di diverse comunità di fitoplancton nella natura.
Smith (1983), dopo aver analizzato la situazione in 12 laghi del mondo, trovò il dominio dei cianobatteri durante i periodi in cui il rapporto azoto/fosforo aveva valori inferiori a 25, con rapporti N/P superiori a 25 predominavano le alghe verdi e diatomee, questo rapporto è stato aumentato a causa della rimozione di acque reflue fosforose. Tilman (1982) definisce i risultati rivelati da Smith “un impatto drammatico del rapporto N/P sulla composizione tassonomica delle algocenosi del lago”.
Successivamente Smith (1986) ha riassunto i materiali di 22 laghi e ha preso in considerazione le condizioni di luce e la morfometria dei leghi. L’inclusione di questi fattori in aggiunta al rapporto tra azoto e fosforo non ha modificato la conclusione relativa all’impatto significativo del rapporto degli elementi biogenici sulla struttura delle specie del plancton.
A un livello di luce fisso, la biomassa relativa blu-verde aumenta con la diminuzione della disponibilità di luce, ad un livello di luce fisso, la biomassa relativa blu-verde aumenta anche quando diminuisce il rapporto N/P.
Schindler (1977) per molti anni ha condotto uno studio sperimentale su piccoli laghi fertilizzati. In uno dei laghi, alimentato per 6 anni tramite fertilizzanti con il rapporto atomico N/P=30, le alghe del genere Scenedesmus hanno predominavano il plancton durante l’esperimento. Dopo che la proporzione da azoto a fosforo nel fertilizzante fu abbassata al valore di 11 il plancton divenne dominato dai cianobatteri, principalmente Aphanizomenon gracile (Lemm.) Elenk.
In un altro lago il fertilizzante è stato alimentato con il rapporto N/P=11 durante l’esperimento, ciò ha portato al dominio dei cianobatteri azoto fissatori del genere Anabaena. Schindler sottolinea che i cianobatteri non solo dominano a bassi rapporti N/P nell’acqua non fertilizzata dei laghi studiati, ma anche quando i fertilizzanti con basso rapporto N/P venivano aggiunti negli stagni.
Quindi, quando il rapporto N/P nel fertilizzante era basso come 5, si è scoperto che l’aumento della biomassa fitoplanctonica nel lago sperimentale (di un fattore da 4 a 8 rispetto a un lago non fertilizzato) è avvenuto quasi interamente a scapito dei cianobatteri, principalmente del genere Anabaena (Findley e Kasian , 1987).
Pick and Lean (1987), rivedendo una serie di studi sull’influenza di diversi nutrienti sui cianobatteri, ha concluso che per questo gruppo di alghe, oltre alle alte temperature, alla stabilità relativa dello strato d’acqua e alle concentrazioni di carbonio e ferro, con valori del rapporto N/P>30 i cianobatteri diventano rari.
La dipendenza dell’abbondanza di cianobatteri su bassi rapporti N/P e una sufficiente fornitura di fosforo è stata ottenuta anche durante uno studio sul fitoplancton fiorente nel lago Kennedy (Stockner and Shortreed, 1988).
Studi nel lago Soyang in Corea, dove il ruolo dei cianobatteri è aumentato dopo un graduale aumento della concentrazione di fosforo nel 1984-1989 (con un alto contenuto di azoto) e la corrispondente diminuzione del rapporto N/P da 100 a 50, ha portato ad un conclusione simile (Cho et al. , 1990).
Un effetto inverso è stato ottenuto nel sequestro ipertrofico sudafricano (Haarhoff et al., 1992): le misure finalizzate allo sgombero delle acque reflue industriali hanno portato ad un aumento del rapporto N/P nell’acqua da 4 a 25 in moli, dopo che l’intera biomassa di fitoplancton si è abbassata i cianobatteri precedentemente dominanti ( Microcystis aeruginosa ) sono stati sostituiti da alghe verdi. Una dipendenza inversa tra il numero di cianobatteri e il valore del rapporto N/P negli ecosistemi dei laghi finlandesi è stata anche dimostrata utilizzando l’analisi di correlazione statistica di Varis (1991).
Finora ci siamo occupati dell’impatto delle forme minerali di azoto e fosforo sulla struttura del fitoplancton; tuttavia, secondo McQueen e Lean (1987), l’aumento della percentuale annuale di cianobatteri può essere indotto dalla diminuzione del rapporto tra l’azoto nitrato e la quantità totale di fosforo. Viceversa, quando questo rapporto era maggiore di 5, non si osservava mai uno sviluppo di massa di cianobatteri.
Ci sono alcuni dati (Blomqvist et al. , 1989; Klapwijk , 1990) che indicano una profonda ristrutturazione delle algocenosi come risultato di un rapporto N/P modificato nell’acqua. Quindi, quando il lago acido svedese Njupfatet (l’abbondanza generale di fondo di azoto e fosforo erano, rispettivamente, 20/0.4 mg/l, N/P=50) è stato alimentato con 10 mg/l di azoto e 1 mg/l di fosforo (rapporto N/P=10), ha determinato una significativa crescita della biomassa di Merismopedia tenuissima Lemm., Peridinium inconspicuum Lemm. e Dictyosphaerium botritella Kom. et Perm., sebbene solo la prima di queste specie fosse stata tra quelle dominanti prima che fossero fatte le aggiunte (Blomqvist et al., 1989).
Il rapporto tra le forme inorganiche di N e P nel bacino del Reno è diminuito drasticamente negli ultimi 45 anni (Klapwijk, 1990). Il risultato è che per questi anni la composizione delle specie di fitoplancton è diventata più povera, diverse specie sono scomparse.
L’interazione congiunta di N, P e Si può esercitare un’influenza anche sulla composizione di una algocenosi naturale. I cambiamenti a lungo termine nel delta del Reno, espressi nella diminuzione dei rapporti Si/N e Si/P, promuovevano uno sviluppo di massa dei dinoflagellati e dei cianobatteri (Admiraal e Vlugt, 1990), ci si può sbarazzare di questo fenomeno a scapito del carico ridotto di azoto e fosforo.
Shamess et al. (1990) giunsero alla conclusione che la fioritura del fitoplancton poteva essere controllata non solo dalla relazione tra azoto, fosforo e silicio, ma anche da altre sostanze e persino dagli oligoelementi Nei loro esperimenti i rapporti di concentrazione molare di solfato ed il molibdeno giocavano un ruolo dominante nella comunità dei cianobatteri che fissano l’azoto. Con ciò, come sottolineano gli autori, le concentrazioni di questi ioni presi separatamente non hanno influenzato il processo di fissaggio dell’azoto.
Fitoplancton naturale in situ – dati da nuovi esperimenti
Abbiamo studiato l’impatto del rapporto N/P sulla composizione del fitoplancton naturale per tre anni negli stagni di allevamento del pesce nel delta del fiume Volga (Levich e Bulgakov 1992, Levich et al., 1996). L’area degli stagni è di 2÷3 ettari, la profondità è di circa 1,5 m. Da aprile a settembre sono stati forniti fosforo sotto forma di Ca(H2PO4)2 ed azoto sotto forma di NH4+ e NO3- in alcune proporzioni secondo due programmi, sperimentale e di controllo.
Negli stagni di controllo le dosi massime di fertilizzanti erano uguali per tutta la stagione e corrispondevano a 2 mg/l di N e 0,5 mg/l di P (N/P=4) nell’acqua. I nutrienti sono stati forniti in vasche sperimentali a N/P=25÷50/1 durante la maggior parte della stagione (le singole dosi per N variavano da 0,24÷0,95 mg/l durante la stagione, quelle per P variavano da 0,01÷0,03 mg/l).
Ca(H2PO4)2 e NH4+ e NO3- sono stati aggiunti simultaneamente negli stagni da una barca, ma non sono stati mescolati e sono stati distribuiti uniformemente su tutta l’area del laghetto.
Campioni di fitoplancton sono stati prelevati da aprile a settembre in tutti gli stagni una volta ogni 10 giorni, nelle stesse date sono state valutate le concentrazioni di N e P, i campioni sono stati ottenuti da una profondità di 1 m. I metodi di stima della biomassa fitoplanctonica, così come i metodi di misurazione della concentrazione di nutrienti, erano simili ai metodi degli esperimenti in vitro.
A partire da giugno, cioè immediatamente dopo un aumento significativo del valore del rapporto N/P rispetto a quello sotto controllo, nell’esperimento è stata osservata una maggiore biomassa di Clorococcales in contrasto con il controllo. Nel 1988 l’effetto stimolante della fertilizzazione sperimentale per i clorococcalli fu osservato più tardi, in agosto e settembre, pertanto la sua biomassa media della stagione era inferiore nell’esperimento. L’aumento della biomassa dei Chlorococcales negli stagni sperimentali rispetto a quelli di controllo nella seconda metà della stagione è mostrato nella Figura 4A.
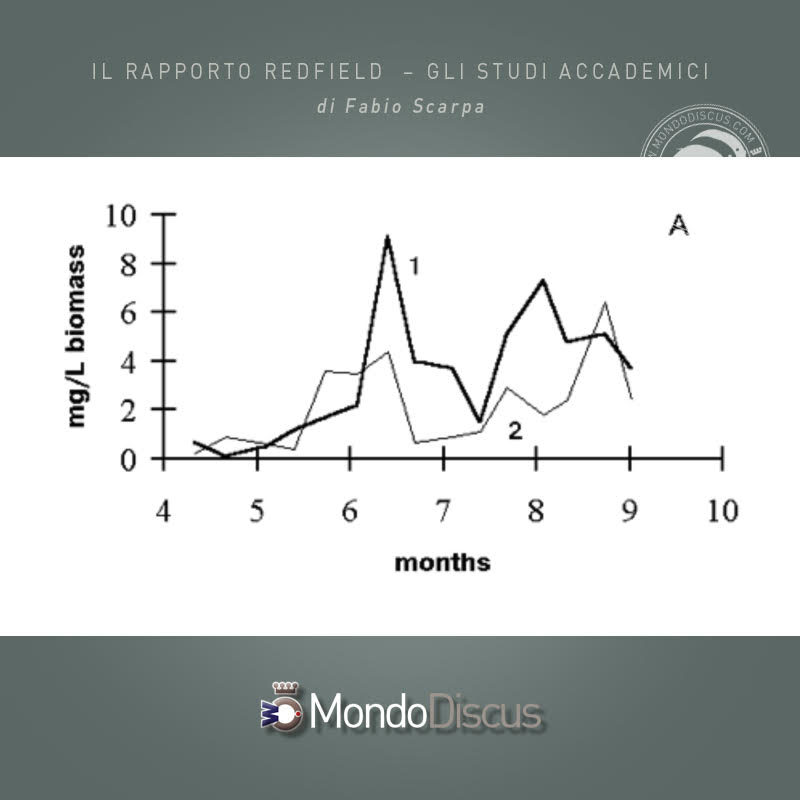
Contemporaneamente, la percentuale dei clorococcalli nella biomassa totale degli stagni sperimentali (con l’eccezione del 1988) è aumentata, a volte la frazione di biomassa cellulare di questo ordine ha raggiunto il 60%, Figura 4B).

Tra le specie dominanti dei Chlorococcales il più sensibile ad un aumento del rapporto N/P era lo Scenedesmus quadricauda. La stimolazione della crescita di Chlorococcales ha portato ad una crescita della biomassa dell’intera divisione Chlorophyta, osservata nella seconda metà della stagione.
Il rapporto N/P aumentato più spesso del solito non ha causato una crescita migliorata di Bacillariophyta e Euglenophyta, per quanto riguarda la biomassa totale di queste divisioni.
Tuttavia, la specie di diatomea dominante, Melosira sp., aumentò la sua biomassa negli stagni sperimentali durante la maggior parte del periodo delle due stagioni (1988-89), nel 1987 questa specie non apparteneva a quelli dominanti.
Un risultato costante della fertilizzazione negli stagni sperimentali era la biomassa soppressa e l’abbondanza frazionaria ridotta dei cianobatteri, a partire dalla metà della stagione (Figura 5).


Questa soppressione è connessa con la diminuzione della biomassa dei generi dominanti di cianobatteri nell’esperimento, cioè Merismopedia e Phormidium, che si è svolta nel periodo da giugno a settembre. I cambiamenti nella struttura dimensionale dell’algocenosi sono stati indicati da una maggiore biomassa di cellule di medie dimensioni, appartenenti alle classi 0,3÷1 e 1÷3,2 ng, in stagni con un rapporto N/P aumentato.
La gestione delle algocenosi dello stagno mediante la regolazione della sua composizione con apporti di elementi nutritivi biogenici è stata effettuata nell’ambito di un programma sperimentale speciale che comprendeva la fertilizzazione degli stagni per l’allevamento di pesci al fine di ottenere una policoltura con prevalenza di specie erbivore. Il programma di fertilizzazione è stato progettato per aumentare sia la biomassa totale del fitoplancton che la % di Clorococcales, il prodotto alimentare più preferibile per i pesci erbivori, sia per diminuire quello di Cyanophyta.
Oltre alle differenze nel rapporto N/P, il sistema di fertilizzazione sperimentale includeva:
1) fornitura più frequente di sostanze minerali (una volta ogni 3 o 4 giorni invece di un programma standard una volta ogni 10 giorni);
2) all’inizio della stagione inizio della fecondazione
3) con dosi iniziali più elevate di fertilizzanti;
4) basso rapporto N/P=4/1 nei fertilizzanti durante aprile-giugno (aprile-maggio 1989);
5) dinamiche stagionali non ritmiche della loro offerta adattate al fabbisogno alimentare avanzato di pesci e zooplancton;
6) ridotte dosi di P e aumentate di N.
Le differenze nei programmi di fertilizzazione tra bacini sperimentali e di controllo sono mostrate nella Tabella 1.

Il programma sperimentale utilizzato in questi esperimenti ha permesso di aumentare la produzione di pesci erbivori del 30% e per ridurre il consumo incompleto di P da parte delle piante acquatiche con la contemporanea conservazione dei livelli normali di N e P nei tessuti di pesce.
Discussione
Come è stato mostrato sopra, le variazioni dei rapporti di nutrienti biogenici nel mezzo hanno influenzato le composizioni di specie e dimensioni delle comunità di fitoplancton, tuttavia, il ruolo di questo fattore rispetto ad altri parametri, come le variazioni nelle concentrazioni assolute di N e P, era aperto alla domanda. I risultati dei nostri esperimenti negli stagni descritti nel capitolo precedente (Levich e Bulgakov 1992) hanno facilitato la sua analisi. Va ricordato che, sebbene i cambiamenti nella struttura del fitoplancton siano stati monitorati durante il periodo di osservazione di tre anni (1987÷1989), nessuno dei componenti del programma di fertilizzazione sperimentale (con l’eccezione del rapporto N/P migliorato) è stato mantenuto costante.
Nel 1987, tali componenti del sistema di fertilizzazione come applicazione di una dose iniziale potenziata e di una più frequente fornitura di sostanze nutritive furono eliminati, ma il fenomeno della regolazione della struttura della comunità algale rimase.
Le concentrazioni assolute dei fertilizzanti forniti nelle vasche sperimentali e di controllo variavano durante tutto il periodo di osservazione, mentre l’effetto del riarrangiamento della struttura del fitoplancton si manifestava dopo la metà dell’anno, cioè immediatamente dopo l’aumento del rapporto N/P. Allo stesso tempo, l’assenza nel 1989 di componenti come il basso rapporto N/P=4/1 nei fertilizzanti introdotti negli stagni in primavera ha portato a una situazione in cui la ridistribuzione della biomassa Cyanophyta si è verificata già all’inizio del Giugno. Tuttavia, la ridistribuzione della biomassa non è stata osservata prima della fine di giugno-luglio del 1987 e 1988, quando il basso rapporto N/P=4/1 ha influenzato la crescita del fitoplancton per un tempo sufficientemente lungo dopo la fecondazione.
Questi dati suggeriscono che la biomassa relativa di taxon e gruppi di dimensioni dipendeva in gran parte dalle concentrazioni relative di N e P e non dall’aumento della concentrazione di N nell’acqua. Risultati simili sono stati riportati da altri autori (Holm & Armstrong 1981).
È noto che a bassi rapporti N/P nell’acqua dello stagno e limitazione di N, le alghe blu-verdi che assimilano N dall’aria atmosferica traggono vantaggio rispetto ai rappresentanti di altri taxa. Così, nei laghi ipertrofici la minimizzazione del rapporto N/P precedeva la fioritura di N fissatori (Blomqvist et al., 1990).
Viceversa, l’aumento dell’ingresso N nel lago ha inibito la crescita del fitoplancton, in cui blu-verde Aphanizomenon flos-aquae è risultata predominante (Elizarova e Korolyov 1990). È noto, tuttavia, che le alghe blu-verdi possiedono proprietà di fissazione dell’N2.atmosferico, questa capacità è una caratteristica dei cianobatteri eterocitosi (Bothe 1982, rappresentanti dei generi Anabaena e Aphanizomenon. Alcune specie di alghe non eterocitose (ad es. Oscillatoria ) possono anche fissare N (Carpenter & Price 1976, Bryceson & Fay 1981). Nei nostri studi, questi generi non erano mai predominanti nella biomassa (Levich e Bulgakov 1992).
La differenza tra il controllo e gli stagni sperimentali è stata determinata dal declino delle biomasse di generi di Merismopedia e Phormidium (in alcuni casi, Aphanothece e Microcystis ) privi di capacità di fissazione dell’N2 . Secondo altri autori (Varis 1992), l’offerta limitata di N è favorevole sia per le alghe blu-verdi che per i rappresentanti del genere Microcystis, pertanto, i verdi blu, come altri taxa algali, rispondono più intensamente alle quantità relative di elementi biogeni rispetto alle loro concentrazioni assolute.
Le differenze nelle risposte dei diversi rappresentanti di una comunità di microalghe ai cambiamenti nel rapporto di fornitura delle risorse nell’ambiente sono determinate da meccanismi fisiologici adattivi che agiscono a livello cellulare. Tale meccanismo è descritto dal concetto di fabbisogno di fitoplancton per componenti di nutrizione minerale (Droop 1973, Rhee 1978, Tilman 1982, Sommer 1983, Levich 1989, Levich et al., 1997). Con questo concetto, il requisito di una specie per N, P, ecc. è uguale alla diminuzione dell’elemento corrispondente nell’ambiente relativo a una singola cellula o ad una quantità unitaria di biomassa. In altre parole, dipende dalla presenza di una sostanza assimilata in una cellula (una quota) se la respirazione, l’escrezione, ecc. vengono trascurate.
Una quota è una quantità specifica per una specie, per una singola specie può variare tra i valori massimi e minimi nel processo di sviluppo della popolazione. La quota minima è una quantità minima di un substrato all’interno di una cella che è ancora in grado di dividere. Secondo Droop (1973), nella curva di crescita logistica della crescita del lotto, una quota minima raggiunta significa che è stata raggiunta una fase stazionaria. Si è tentati di concludere che se una certa specie possiede un certo rapporto di quote minime di N e P, ad esempio, questo rapporto indica il grado della sua competitività a uno o ad un altro valore del rapporto N/P nell’acqua.
Il rapporto di concentrazione dell’elemento biogenico, più vicino al rapporto tra le quote minime, è ottimale per una data specie.
Idee simili sono state avanzate da Rhee & Gotham (1980) che hanno trovato rapporti N/P ottimali per nove specie microalgali e hanno sottolineato che questi rapporti corrispondono ai rapporti tra i requisiti di questi elementi.
Nell’ambito del modello variazionale di una comunità di fitoplancton (Levich 1980), il teorema della massima abbondanza di specie è stato dimostrato utilizzando modelli matematici (Levich et al 1993, Alexeyev, Levich 1997). Secondo questo teorema, l’abbondanza relativa delle specie dipende esclusivamente dai rapporti delle risorse ambientali che limitano la crescita della comunità. I coefficienti di risorse che forniscono la quota percentuale massima di una specie in una comunità sono uguali ai rapporti delle quote minime di cellule per questa specie particolare (Droop 1973). Di fatto, l’ipotesi conclusa nella formulazione del teorema sul massimo delle abbondanze di specie ci ha stimolato a condurre una serie di esperimenti e cercare dati letterari più recenti riguardanti il ruolo dei rapporti nutrizionali nella struttura delle comunità di specie di algocenosi.
Tuttavia, per le comunità multispecie, la dipendenza dei risultati della competizione per le risorse sul gradiente dei rapporti delle risorse consumate, sono anche adeguatamente descritte da altri modelli costruiti con l’uso di diversi approcci metodologici. Pertanto, la teoria grafica della competizione delle risorse proposta da Tilman (1982) consente di prevedere l’abbondanza di specie coesistenti nelle comunità in base a rapporti di risorse medie e requisiti di specie in queste risorse (secondo Tilman, il termine “requisiti di specie” è applicato a una risorsa consumata da una popolazione nella fase stazionaria della sua crescita quando il suo incremento è esattamente uguale alla mortalità all’interno della popolazione). L’analisi di tale competizione con l’aiuto di equazioni differenziali classiche del tipo Lotka e Volterra ha dimostrato (vedi anche Abrosov 1977) che le aree di coesistenza o eliminazione di specie in una comunità sono divise da valori soglia di rapporti di risorse in un mezzo. Un’analisi diretta computerizzata che utilizzava un modello fenomenologico di algocenosi simulando la dinamica delle concentrazioni intracellulari di N e P (Jorgensen 1980), ha rivelato un’evidente alterazione della classificazione delle specie con un cambiamento nel rapporto N: P (Levich et al., 1997).
I riferimenti a un rapporto “ottimale” di risorse nutritive possono essere trovati in numerose relazioni di altri ricercatori impegnati nello studio di comunità di laboratorio e di fitoplancton naturale, molte di queste osservazioni sono state descritte nelle sezioni precedenti di questa recensione.
Tilman (1982) richiama l’attenzione sulle sorprendenti coincidenze tra rapporti di risorse ottimali per singole specie determinate in condizioni di laboratorio e quelle in cui alcune specie prevalgono nelle comunità naturali. In particolare, citando le opere di Smith (1983) che ha studiato l’influenza del rapporto N/P sul predominio delle alghe blu-verdi nei laghi del mondo, Tilman lo ha definito “drammatico” per la composizione tassonomica del fitoplancton lacustre. Va ricordato a questo proposito che negli studi empirici il termine “specie ottimale” è usato per definire un rapporto risorsa, la cui approssimazione fa aumentare la sua biomassa ad una data specie.
Il suddetto teorema che postula un massimo di abbondanze di specie definisce più concretamente le condizioni dei criteri di ottimalità: il rapporto ottimale di risorse è associato alle caratteristiche fisiologiche specifiche delle specie di organismi, come le quote cellulari di queste risorse nel caso del fitoplancton, e assicura la massima biomassa relativa di una specie in una comunità.
Va sottolineato che la formulazione dell’ipotesi che postula il contributo dei rapporti di risorse alla struttura comunitaria non diminuisce il ruolo delle concentrazioni assolute delle risorse disponibili per questa comunità, mentre le concentrazioni assolute di risorse medie determinano la biomassa totale di una comunità, i rapporti di queste risorse determinano la biomassa relativa delle singole specie nella comunità.


| Il Rapporto Redfield: la pratica | |
| Il Rapporto Redfield: la pratica | |